大家好,这里是专注表观组学十余年,领跑多组学科研服务的易基因。
组蛋白及其翻译后修饰在调控基因表达和其他染色质模板化过程中起着重要作用。组蛋白H3赖氨酸4三甲基化(H3K4me3)和组蛋白H3K27me3分别在小鼠植入前胚胎的基因激活和基因抑制中发挥重要作用。H3K4me3和H3K27me3组成的染色质区域称为二价结构域,在小鼠胚胎干细胞(mESC)中普遍存在。由二价结构域标记的基因通常处于蓄势状态,可迅速激活以响应发育线索。多梳抑制复合体2(polycomb repressive complex 2,PRC2)介导染色质H3K27me3(包括二价结构域);而H3K4me3对于阻止这些基因DNA甲基化至关重要。那么二价结构域是如何建立和调控的呢?
2023年05月27日,复旦大学生物医学研究院/上海市医学表观遗传学重点实验室沈宏杰/徐文绮和牛津大学yang shi教授合作以“METTL14 regulates chromatin bivalent domains in mouse embryonic stem cells”为题在《cell reports》期刊发表研究论文,该研究以小鼠胚胎干细胞(mouse embryonic stem cell,mESC)为研究对象,通过染色质免疫共沉淀测序(ChIP-seq)、RNA-seq等研究揭示了METTL14在调控二价结构域和基因表达中的作用。

标题:METTL14 regulates chromatin bivalent domains in mouse embryonic stem cells(METTL14调控小鼠胚胎干细胞中的染色质二价结构域)
时间:2023-05-27
期刊:cell reports
影响因子:IF 9.995
技术平台:ChIP-seq、RNA-seq、Western blot、qRT-PCR等
摘要:
METTL14(methyltransferase-like 14)是一种RNA结合蛋白,可与METTL3结合以介导N6-甲基腺苷(m6A)甲基化。最近的研究表明了METTL3在小鼠胚胎干细胞(mESC)异染色质中的作用,但METTL14在小鼠胚胎干细胞异染色质中的分子功能尚不清楚。本研究分析了METTL14特异性地结合和调控以H3K27me3 H3K4me3为标记的二价结构域。研究结果表明,METTL14基因敲除导致H3K27me3水平降低,以及H3K4me3水平升高导致转录增加。METTL14对二价结构域的调控与METTL3或m6A修饰无关。METTL14通过与H3K27甲基转移酶多梳抑制复合体2(PRC2)和H3K4去甲基化酶KDM5B互作并可能将其招募到染色质,从而增强H3K27me3并减少H3K4me3。本研究揭示了METTL14在维持mESC中二价结构域完整性中的作用,且与METTL3无关,从而表明哺乳动物中二价结构域的调控机制。
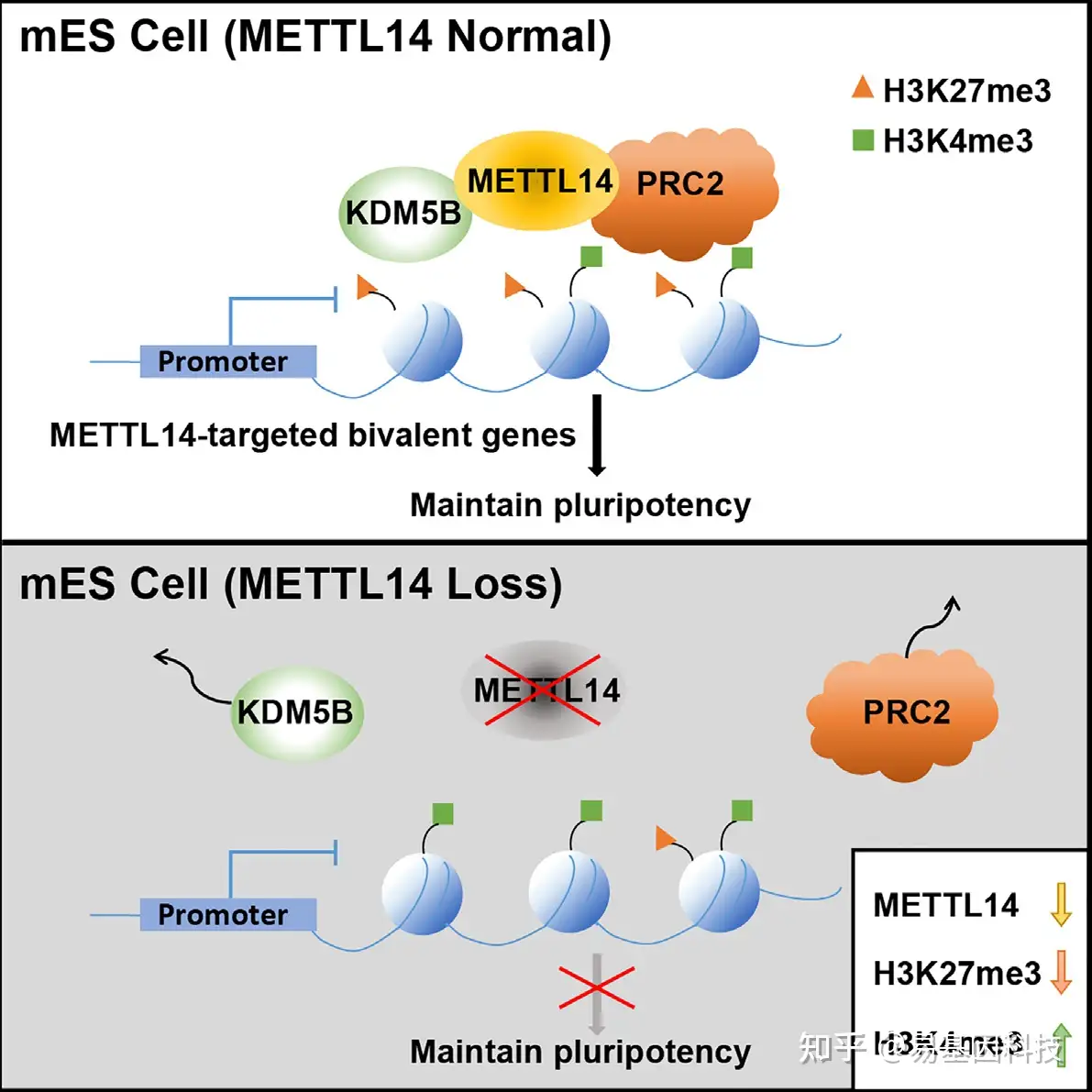
研究图形摘要
研究要点:
- METTL14主要在小鼠胚胎干细胞的二价结构域中富集
- METTL14缺失导致二价结构域中的H3K27me3减少和H3K4me3增加
- METTL14调控二价结构域,与METTL3或m6A修饰无关
- METTL14可能招募H3K27me3甲基转移酶PRC2和H3K4me3去甲基化酶KDM5B
研究结果:
- ChIP-seq揭示METTL14主要与二价结构域结合
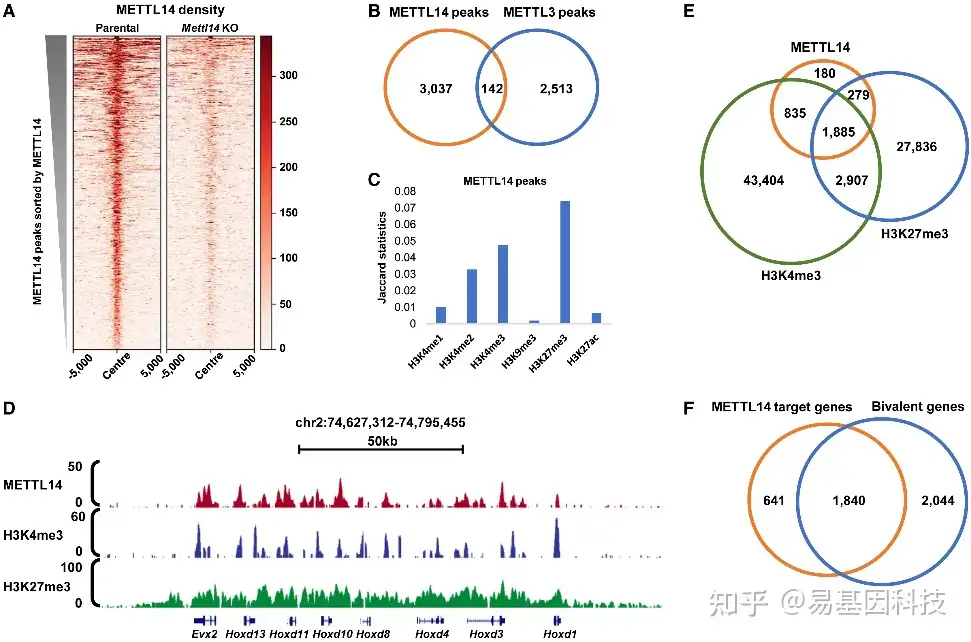
图1:METTL14与二价结构域结合
(A)亲本细胞系和METTL14-KO细胞系中METTL14 peaks(n=3179)的METTL14水平分布热图。
(B) METTL14和METTL3 peaks之间重叠的Venn图。p=1.24e−175,Fisher精确检验。
(C) METTL14 peaks与不同组蛋白修饰peaks重叠率条形图(Jaccard统计)。
(D) UCSC基因组浏览器快照显示METTL14和组蛋白修饰在代表性二价结构域的结合模式。
(E) METTL14 peaks和二价结构域之间的重叠Venn图。p=0,Fisher精确检验。
(F) METTL14靶基因和二价基因之间的重叠Venn图。p=0,Fisher精确检验。
*热图根据(A)中亲本细胞中METTL14水平的降序排列。H3K4me3和H3K27me3 ChIP-seq两个重复。
(2)METTL14 缺失导致二价结构域的H3K27me3降低和H3K4me3水平升高
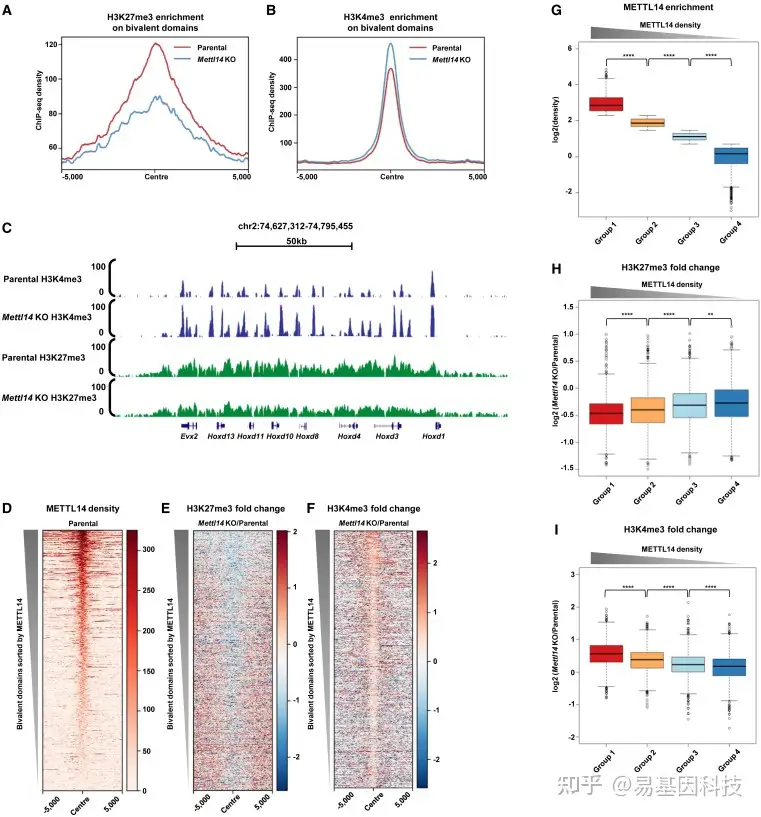
图2:METTL14调控二价结构域的组蛋白修饰
(A-B)亲本细胞系和Mettl14-KO细胞系中二价结构域(n=4863)处H3K27me3(A)和H3K4me3(B)的平均富集聚合图。
(C) UCSC基因组浏览器快照显示亲本细胞系和Mettl14-KO细胞系中代表性二价结构域上组蛋白修饰的结合模式。
(D)亲本细胞系中二价结构域(n=4863)的METTL14水平热图。
(E-F)H3K27me3(E)和H3K4me3(F)在二价结构域(n=4863)的ChIP-seq倍数变化(log2(Mettl14 KO/亲本))热图。
(G) METTL14在二价结构域的水平(log2(density))箱形图,根据METTL14结合水平分布从高到低分为四组(1、2、3和4组分别n=1216、1216、1216和1215个peaks)。∗∗p<0.0001(从左到右的精确p值:0、0和0),双侧配对t检验。
(H-I)H3K27me3(H)和H3K4me3(I)在四组二价结构域上的ChIP-seq倍数变化箱形图(log2(Mettl14 KO/亲本)),如(G)所示p<0.01,****p<0.0001(H从左到右的精确p值:2.34e-06、2.12e-09和8.60e-03;I从左到右的精确p值:3.45e-30、1.52e-16和1.19e-07),双侧t检验。
(3)METTL14缺失导致二价基因转录增加
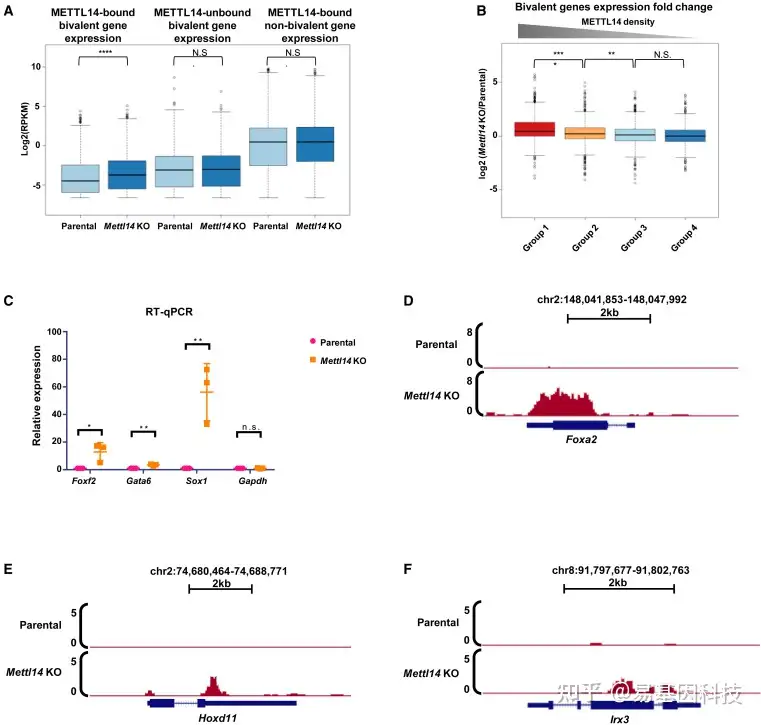
图3:METTL14调控二价基因的转录
(A) 亲本细胞系和METTL14-KO细胞系中METTL14结合二价基因(n=1840),METTL14未结合二价基因(n=2044)和其他METTL14结合基因(n=641)的RNA-seq基因表达箱形图(log2(RPKM))p<0.0001(从左到右的精确p值:8.08-10、3.35e-01和5.09e-01),双侧t检验。
(B) 与图2G中的四组二价结构域相关的二价基因RNA-seq基因表达倍数变化箱形图(log2(Mettl14-KO/亲本))(1、2、3和4组分别n=1087、1035、973和789个基因)。**p<0.01,****p<0.0001(从左到右的精确p值:2.19e–14、5.14e–03和3.25e–01),双侧t检验。
(C) qRT-PCR分析亲本细胞系和Mettl14-KO细胞系中二价基因(Foxa2、Irx3、Hoxd11)的相对表达水平。显示了三个生物学重复的平均值±SDp<0.05,**p<0.01(从左到右的精确p值:3.69e–02、2.50e–03、9.81e–03和9.70e–01),双侧t检验。
(D–F) UCSC基因组浏览器快照显示了亲本细胞系和Mettl14-KO细胞系中二价基因(Foxa2、Irx3、Hoxd11)的转录水平。
(4)METTL14调控二价结构域,与METTL3无关
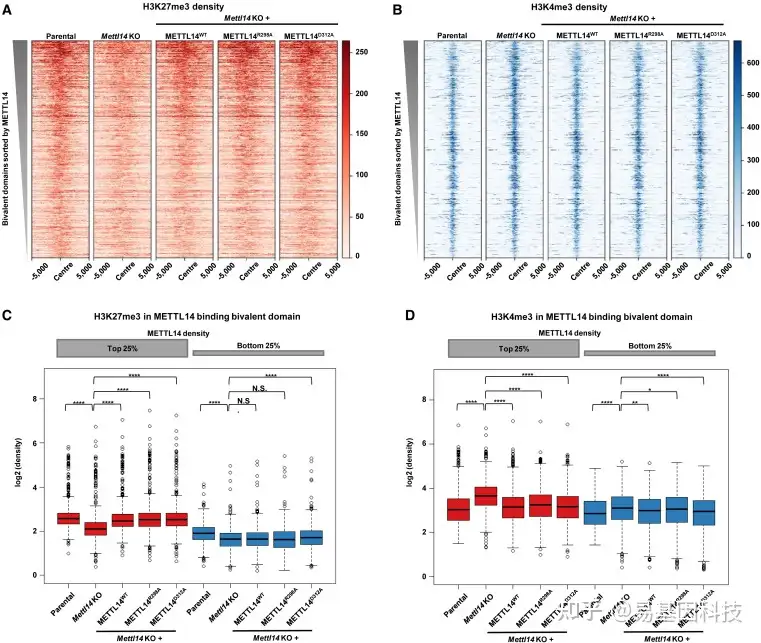
图4:METTL14调控二价结构域与m6A无关
(A-B) 亲本细胞系、Mettl14-KO细胞系和含有METTL14-WT、METTL14-R298A或METTL14-D312A的挽救细胞系的二价结构域(n=4863)中的H3K27me3(A)和H3K4me3(B)的水平分布。
(C-D) H3K27me3(C)和H3K4me3(D)在二价结构域中第1组和第4组上的水平分布(log2(density)),如图2G所示,在亲本细胞系、Mettl14-KO细胞系和含有METTL14-WT、METTL14-R298A或METTL14-D312A的挽救细胞系中p<0.05,**p<0.01,****p<0.0001(C从左到右的精确p值:1.38e–80、5.18e–52、1.14e–56、3.78e–58、1.08e–44、1.22e–01、6.66e–01和1.68e–05;D从左到右的精确p值:1.67e–78、2.75e–64、4.01e–44、3.61e–61、1.04e–06、1.92e–04、1.28e E–02和8.82e–10),双侧t检验。
(5)METTL14与H3K4me3甲基转移酶复合体PRC2和H3K4me3去甲基化酶KDM5A和KDM5B互作
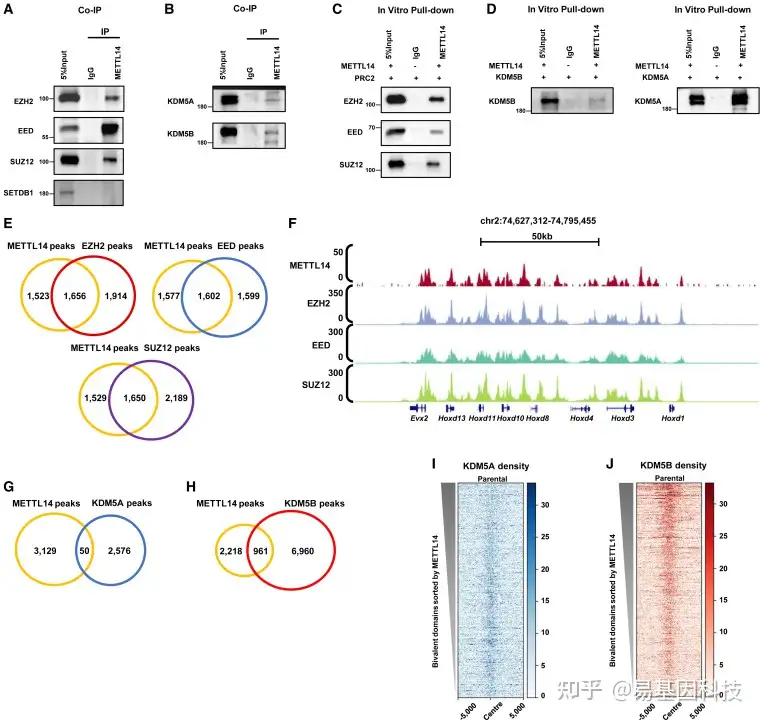
图5:二价结构域中的PRC2和KDM5A/5B定位与METTL14相关
(A) METTL14和PRC2复合体(EZH2、EED、SUZ12)的相互免疫沉淀Western blot实验。
(B) METTL14和KDM5A/KDM5B相互免疫沉淀Western blot实验。
(C) METTL14和PRC2复合体(EZH2、EED、SUZ12)的体外下拉Western blot实验。
(D) METTL14和KDM5A(左)以及KDM5B(右)的体外下拉Western blot实验。
(E) METTL14和EZH2以及EED和SUZ12之间的重叠Venn图(从左到右的精确p值:0、0和0)。Fisher精确检验。
(F) UCSC基因组浏览器快照显示亲本细胞系中METTL14、EZH2、EED和SUZ12在二价结构域的水平分布。
(G-H)METTL14与KDM5A(G)和KDM5B(H)之间的重叠Venn图(G的精确p值:8.07e–41;H的精确p值:0),Fisher精确检验。
(I–J)亲本细胞系中KDM5A(I)和KDM5B(J)在二价结构域(n=4863)中的水平分布热图。
(6)METTL14与PRC2和KDM5B共定位,是PRC2在二价结构域结合所必需的
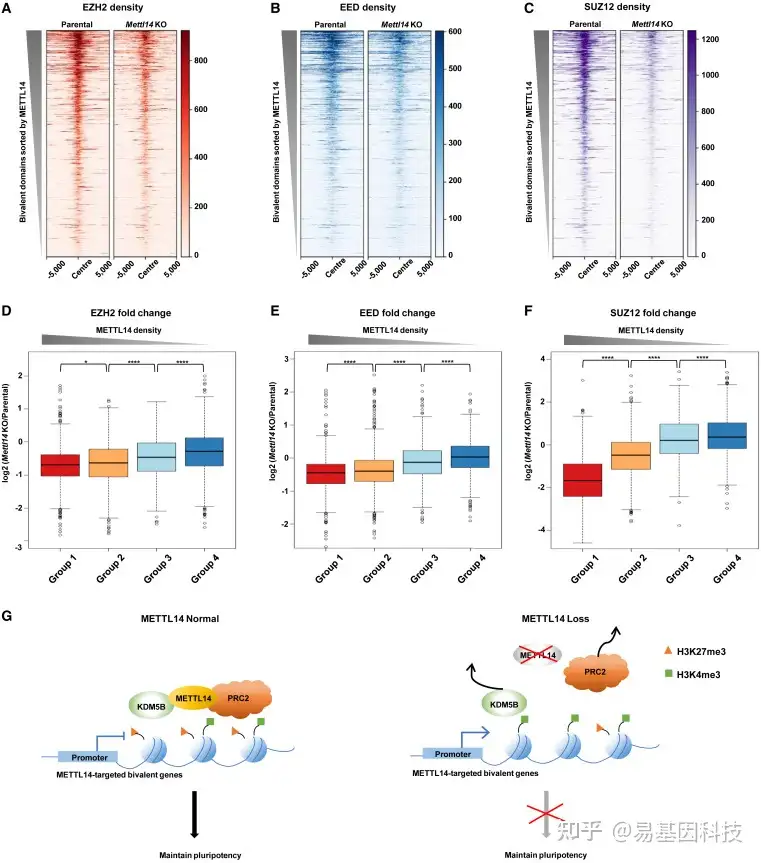
图6:在二价结构域中,METTL14调控PRC2
(A–C) 亲本细胞系和Mettl14-KO细胞系中二价结构域(n=4863)中的EZH2(A)、EED(B)和SUZ12(C)水平分布热图。
(D–F) EZH2(D)、EED(E)和SUZ12(F)在四组二价结构域上的ChIP-seq倍数变化箱形图(log2(Mettl14 KO/亲本)),如图2G所示p<0.05,****p<0.0001(D从左到右的精确p值:1.71e–02、3.50e–13、5.34e–08;E从左到右的精确p值:3.74e–07、3.04e–27和1.80e–12;F从左到右的精确p值:7.19e–145、1.79e–71和6.19e–06),双侧t检验。
(G) METTL14调控PRC2和KDM5B在二价结构域的定位示意图模型。METTL14缺失导致H3K27me3降低和H3K4me3增加以及随后METTL14结合二价基因的激活。
易基因小结
本研究通过ChIP-seq等实验研究,揭示了在小鼠胚胎干细胞(ESC)中,与在重复元件中富集的METTL3不同,METTL14主要富集在由H3K27me3和H3K4me3修饰的基因组区域,即二价结构域。研究结果表明METTL14缺失导致H3K27me3显著性和选择性减少,同时二价结构域中的H3K4me3增加,从而导致基因激活,表明了METTL14对于二价结构域的完整性非常重要。从机制上讲,METTL14通过促进H3K27me3 writers、PRC2复合体和H3K4me3 erasers KDM5B的结合来维持二价结构域。因此,本研究揭示了METTL14在调控二价结构域和基因表达中的作用,与METTL3介导的m6A无关。
关于易基因染色质免疫共沉淀测序 (ChIP-seq)
染色质免疫共沉淀(Chromatin Immunoprecipitation,ChIP),是研究体内蛋白质与DNA相互作用的经典方法。将ChIP与高通量测序技术相结合的ChIP-Seq技术,可在全基因组范围对特定蛋白的DNA结合位点进行高效而准确的筛选与鉴定,为研究的深入开展打下基础。
DNA与蛋白质的相互作用与基因的转录、染色质的空间构型和构象密切相关。运用组蛋白特定修饰的特异性抗体或DNA结合蛋白或转录因子特异性抗体富集与其结合的DNA片段,并进行纯化和文库构建,然后进行高通量测序,通过将获得的数据与参考基因组精确比对,研究人员可获得全基因组范围内某种修饰类型的特定组蛋白或转录因子与基因组DNA序列之间的关系,也可对多个样品进行差异比较。
应用方向:
ChIP 用来在空间上和时间上不同蛋白沿基因或基因组定位
- 转录因子和辅因子结合作用
- 复制因子和 DNA 修复蛋白
- 组蛋白修饰和变异组蛋白
技术优势:
- 物种范围广:细胞、动物组织、植物组织、细菌微生物多物种富集经验;
- 微量建库:只需5ng以上免疫沉淀后的DNA,即可展开测序分析;
- 方案灵活:根据不同的项目需求,选择不同的组蛋白修饰特异性抗体。
技术路线:
易基因提供全面的DNA与蛋白互作测序方案。
参考文献:
Mu M, Li X, Dong L, Wang J, Cai Q, Hu Y, Wang D, Zhao P, Zhang L, Zhang D, Cheng S, Tan L, Wu F, Shi YG, Xu W, Shi Y, Shen H. METTL14 regulates chromatin bivalent domains in mouse embryonic stem cells. Cell Rep. 2023 Jun 13;42(6):112650.
相关阅读:
项目文章 | NAR:ChIP-seq等揭示蛋白质酰基化与c-di-GMP协同调控放线菌发育与抗生素合成机制
项目文章 | 组蛋白ChIP-seq揭示烟粉虱共生菌Hamiltonella调控宿主生殖新机制