大家好,这里是专注表观组学十余年,领跑多组学科研服务的易基因。
目前,六分之一的夫妇患有不孕不育,其中多达一半的病例由男性因素引起,在过去的40年中,精子数量下降了50%。尽管导致精子质量和数量下降的因素知之甚少,但环境-表观基因组相互作用是可能的。暴露于有毒物质、饮食和 BMI 升高与精子表观基因组变化和男性临床人群的生育能力降低有关。然而,这些研究的重点主要集中在DNA甲基化水平上,而对染色质与精子中DNA甲基化之间相关性知之甚少。父系不良环境暴露与不孕不育和不良结果相关,这种作用可能通过精子的组蛋白修饰传递。到目前为止,对男性精子染色质的深入分析还很有限。
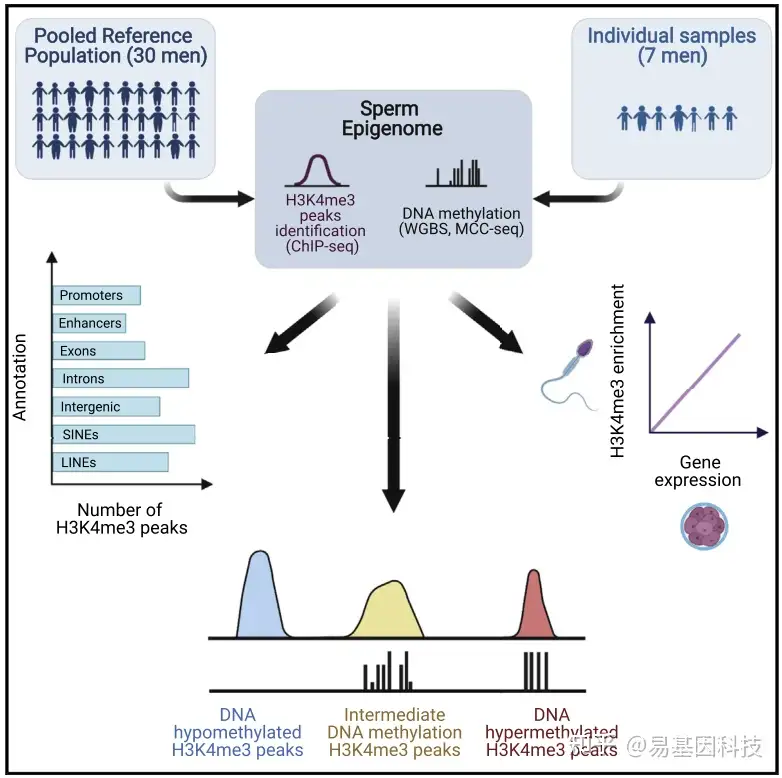
2021年7月20日,加拿大麦吉尔大学Vanessa Dumeaux和Sarah Kimmins团队在《Cell Reports》杂志发表题为“Whole-genome sequencing of H3K4me3 and DNA methylation in human sperm reveals regions of overlap linked to fertility and development”的研究论文,该研究以男性精子样本为对象,通过ChIP-seq和WGBS等技术研究了 H3K4me3 在精子中的位点及其与相同样本中精子 DNA 甲基化谱的关系。

标题:Whole-genome sequencing of H3K4me3 and DNA methylation in human sperm reveals regions of overlap linked to fertility and development(人类精子中H3K4me3和DNA甲基化的全基因组测序揭示了与生育和发育相关的重叠区域)
时间:2021-7-20
期刊:Cell Reports
影响因子:8.8
技术平台:ChIP-seq、WGBS等
研究摘要:
本研究使用深度测序来表征37名男性代表性参考群体中精子的组蛋白H3赖氨酸4三甲基化(H3K4me3)和DNA甲基化图谱。分析结果表明,H3K4me3靶向全基因组和生育和发育相关基因。在原始生殖细胞、胚胎增强子和短穿插核元件(short-interspersed nuclear elements,SINEs)中逃避表观遗传重编程区域也表现出富集。H3K4me3和DNA甲基化在全基因组中存在显著重叠,表明这些标记之间存在潜在的相互作用,而此前报道的这些标记在精子中互斥。精子中H3K4me3标记区域与胚胎转录组的比较分析表明,父本染色质对胚胎基因表达有影响。
研究要点
- 深度测序鉴定人类精子中携带H3K4me3的区域
- H3K4me3广泛分布于基因组中,在启动子和SINE处富集
- 精子H3K4me3标记发育基因并与胚胎基因表达相关
- 除了DNA低甲基化位点外,H3K4me3还与高甲基化区域重叠
研究摘要
研究结果:
- 组蛋白H3K4me3在精子基因和调控区域富集
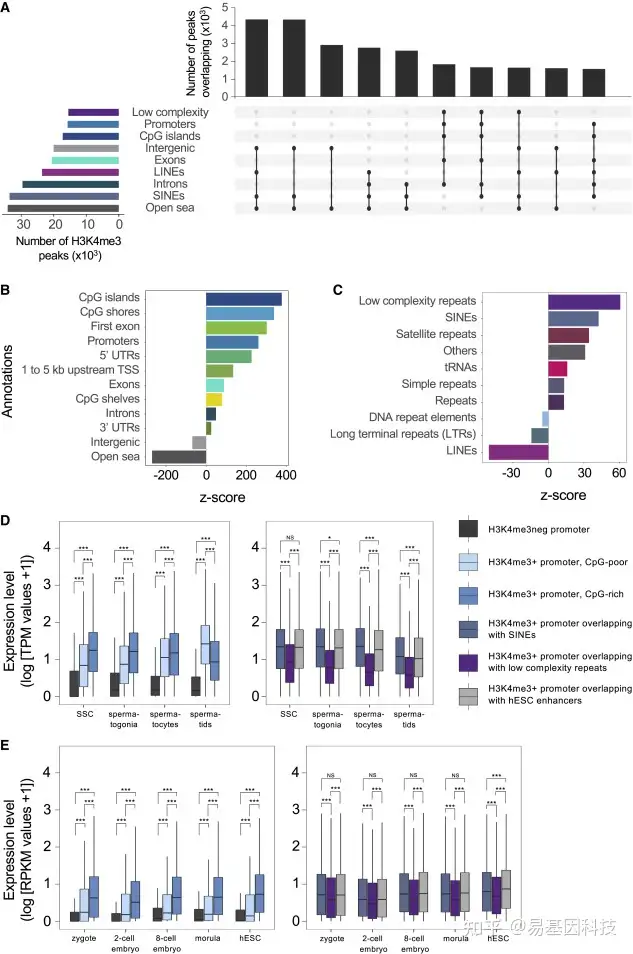
图1:H3K4me3在人类精子的CpG区域、启动子区和SINE区富集
从30名男性的精子中制备样本上进行H3K4me3的ChIP-seq,使用MACS2来命名peaks。
- 基于基因组和CpG注释以及重复元件和低复杂性DNA序列重叠的H3K4me3 peaks位点。从Bioconductor软件包annotatr (Cavalcante and Sartor, 2017)和RepeatMasker (http://www.repeatmasker.org/)中获得注释。不同注释之间peaks重叠通过连接节点来表示,重叠peaks数量显示在它们上方条形图上,显示前10个重叠。
- 每个特定注释的H3K4me3 peaks富集。通过使用Bioconductor封装区域器(Gel et al., 2016)确定Z分数来表示阳性和阴性富集。对于所有显示的注释,通过排列测试,p < 0.001。
- 每次重复注释的H3K4me3 peaks富集;排列检验P < 0.001。
D-E. 箱形图显示精子中启动子不携带H3K4me3的基因表达水平,在位于CpG缺失区域或CpG富集区的启动子上标记了H3K4me3,以及在精子发生(D)和发育©期间启动子与SINEs、低复杂性重复或hESCs增强子重叠。转录水平是基于表达为精原干细胞(SSCs)、精原细胞(有丝分裂期)、精母细胞(减数分裂)和精子细胞(精子发生)中精子发生的TPM转录本丰度或为发育绘制的RPKM片段的千碱基reads来可视化的。P值由成对Wilcoxon秩和检验确定,多重检验采用Benjamini-Hochberg校正。NS表示调整后的p值不显著。*调整后p< 0.05, * * *调整后p < 0.001。
(2)SINE、低复杂性重复序列或胚胎增强子的携带H3K4me3启动子与不同的胚胎过程有关
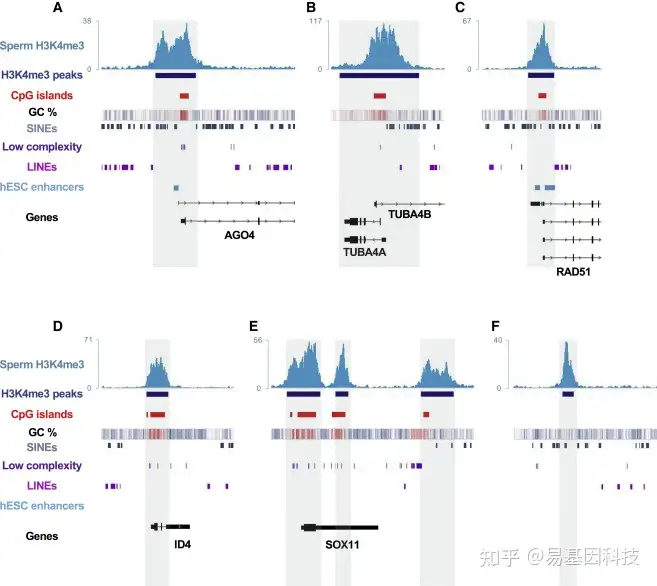
图2:精子H3K4me3 peaks与CpG富集区、重复元件和hESC增强子之间的重叠
(A-F)基因组浏览器快照显示参考基因组的ChIP-seq轨迹(蓝色)、MACS2 calling的H3K4me3 peaks值(深蓝色框)、CpG岛位点轨迹(红色框,UCSC)、GC百分比热图(蓝色,低GC百分比;白色,50%GC;红色,高GC百分比)、SINE位点(深灰色框)、低复杂性重复序列(深紫色框),LINE位点(紫色框)(从RepeatMasker获得)和hESC增强子的位置(浅蓝色框)。H3K4me3与这些元件中的每一个重叠是RNA介导的基因沉默AGO4(A)的Argonaute基因家族成员,编码微管蛋白亚型TUBA4A和TUBA4B(B)的两个基因,辅助双链断裂修复RAD51(C)的基因,发育基因ID4(D)和SOX11(E),以及最后对于随机基因间区域(F)进行了说明。
(3)H3K4me3 和 DNA 甲基化同时发生在人类精子的功能基因组区域
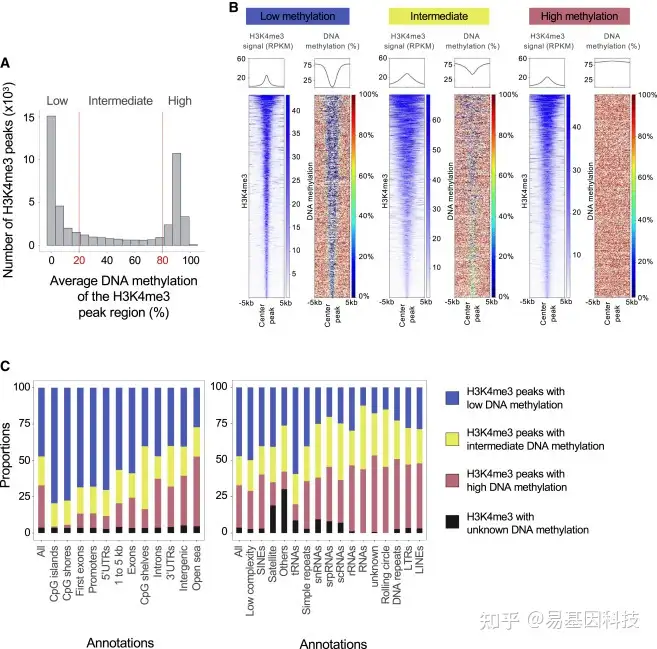
图3:人类精子中H3K4me3 peaks值区域的DNA甲基化水平
对用于H3K4me3 ChIP-seq的30名男性精子进行WGBS,并评估位于H3K4me3 peak内CpGs的DNA甲基化水平。然后计算peaks标记区域的平均DNA甲基化水平。
- 具有低DNA甲基化(低甲基化,甲基化≤20%)、中等DNA甲基化(20%≤甲基化≤80%)或高DNA甲基化(高甲基化,甲基化≥80%)的H3K4me3 peaks数。
- H3K4me3信号(单位:RPKM)中低甲基化(23023个peaks)、中等甲基化(8,087个peaks)或高甲基化(16,777个peaks)H3K4me3 peaks中心(±5kb)的DNA甲基化水平热图(单位:%)。每条线代表一个H3K4me3 peaks。所有peaks的H3K4me3信号和DNA甲基化在热图的顶部总结。
© H3K4me3 peaks内基因组、CpG、RNA和重复序列注释的低甲基化、中等甲基化和高甲基化DNA比例。
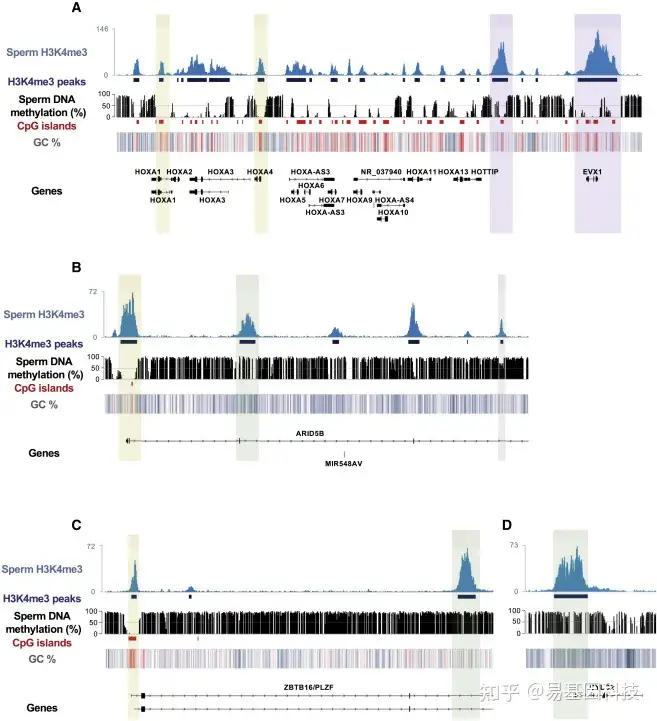
图4:人类精子中H3K4me3和DNA甲基化谱
H3K4me3 peaks和DNA甲基化之间的重叠显示在基因组浏览器快照中,显示了HOXA发育基因簇(A)的ChIP-seq轨迹(蓝色)、MACS2 calling的H3K4me3 peaks(深蓝色)、WGBS轨迹(黑色)、CpG岛位点轨迹(红色,UCSC)和GC百分比热图(蓝色,低GC %;白色,50% GC;红色,高GC %),染色质修饰蛋白ARID5B (B),精原细胞标记物ZBTB16/PLZF (C)和精子发生基因CYLC2 (C)。H3K4me3 peaks位于非DNA甲基化的CpG岛(黄色),包括不同甲基化水平区域(紫色),CpG DNA低甲基化区域(绿色)和中等DNA甲基化区域(灰色)。
(4)精子H3K4me3与DNA甲基化在精子发生和胚胎发生过程中基因调控的功能关系探讨
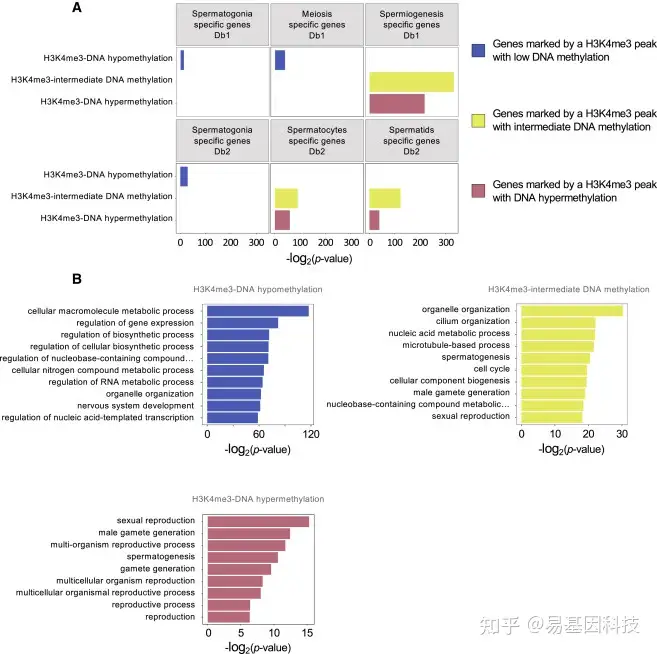
图5:DNA甲基化水平在精子发生、胚胎发生和基本细胞过程的功能基因启动子区H3K4me3 peaks变化
- H3K4me3过表达在精子发生相关基因启动子区域具有低、中、高平均DNA甲基化peaks值。与早期精子发生、减数分裂和精子发生相关的基因选自两个数据库:Db1(Jan et al.,2017)和Db2(Wang et al.,2018)。给出了Benjamini-Hochberg校正的超几何最小似然p值。
- 基因启动子(TSS上游1kb)中的H3K4me3 peaks的GO分析。通过PANTHER过表征测试(GO生物过程完成)检测特定生物过程中的富集度。P值通过Fisher精确检验确定,并对多次检验进行Bonferroni校正。过滤富集度≥1.25的非冗余过程后,显示p值最低的前10个过程。
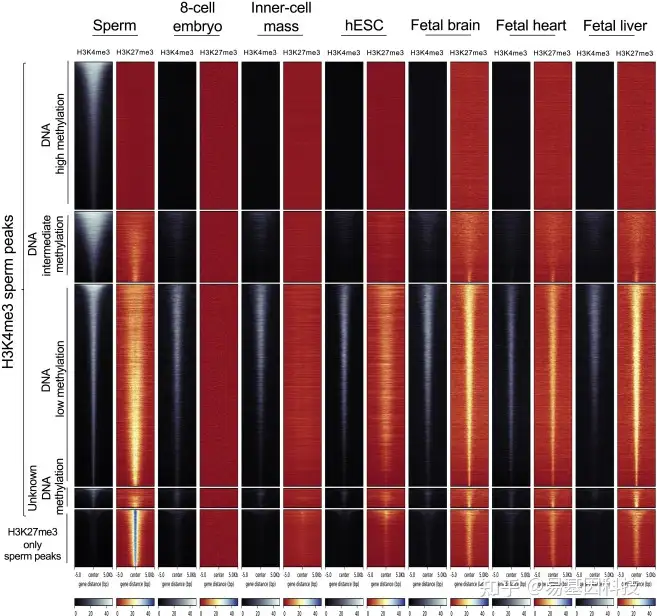
图6:H3K4me3精子peaks区域的H3K4me3/H3K27me3二价性,存在低、中或高DNA甲基化
H3K4me3和H3K27me3信号(单位:RPKM)在H3K4me3 peaks中心(±5kb)在人类精子(参考30名男性的H3K4me3和已发表的H3K27me3数据(hamoud etal ., 2009))、8细胞人类胚胎和内细胞团(Xia etal .,2019)、 hESC (Grandy etal ., 2015)和人类胎儿器官(Yan etal .,2016) 热图。每个通道代表在参考人群中鉴定的具有DNA低甲基化(23,023)、中甲基化(8,087)、高甲基化(16,777)或未知甲基化(没有CpG或在WGBS中覆盖小于10倍的CpG)的H3K4me3 peaks区域,或者未被H3K4me3标记的H3K27me3精子peaks区域。
(5)精子中的H3K4me3靶向增强子和逃避表观遗传重编程区域
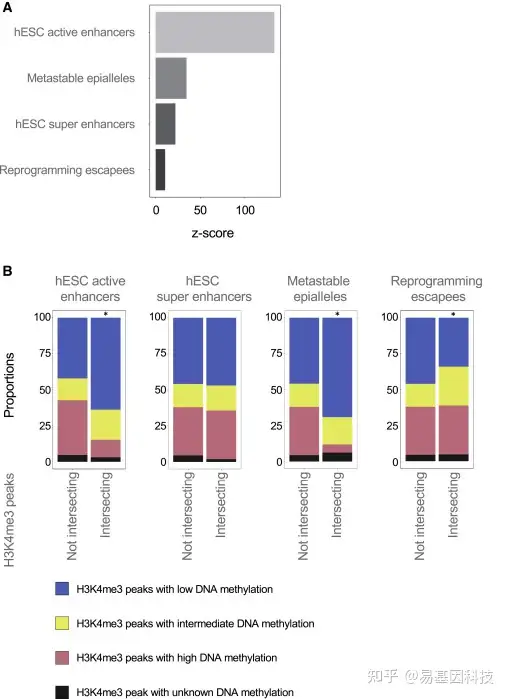
图7:H3K4me3在hESC增强子和逃避重编程区域富集
(A) H3K4me3富集在已鉴定的hESC增强子和超级增强子处达到峰值(Barakat等人,2018),在亚稳态表等位基因区域(Kessler et al.,2018)以及在人类原始生殖细胞(PGCs)中通过DNA甲基化缺失而逃避重编程区域达到峰值(Tang等人,2015)。正富集Bioconductor包regioneR确定的Z评分表示。对于显示的所有区域,通过排列检验,p<0.001。
(B) 与感兴趣区域相交或不相交的低甲基化、中等甲基化和高甲基化H3K4me3 peaks比例。不相交于感兴趣区域的peaks和相交的peaks比例变化通过χ2检验进行检验,星号表示p<0.001。
本研究的局限性
本研究遵循ENCODE指南,使用DNA甲基化分析的金标准——WGBS,和H3K4me3的深度ChIP-seq分析了加拿大男性代表群体的精子表观基因组。ChIP-seq在7名男性个体中进行验证,WGBS通过定制精子MCC-seq方法进行验证。这种方法的局限性是MCC-seq不能覆盖全基因组。但本研究捕获中覆盖区域验证了WGBS的区域。当然有可能忽略到某些区域的H3K4me3和DNA甲基化存在很强的个体变异性。在未来的研究中,尽管组蛋白保留水平低,但一旦染色质的单细胞分析足够敏感,可以分析精子,就可以解决精子与精子之间的异质性。
关于易基因全基因组重亚硫酸盐测序(WGBS)
全基因组重亚硫酸盐甲基化测序(WGBS)可以在全基因组范围内精确的检测所有单个胞嘧啶碱基(C碱基)的甲基化水平,是DNA甲基化研究的金标准。WGBS能为基因组DNA甲基化时空特异性修饰的研究提供重要技术支持,能广泛应用在个体发育、衰老和疾病等生命过程的机制研究中,也是各物种甲基化图谱研究的首选方法。
易基因全基因组甲基化测序技术通过T4-DNA连接酶,在超声波打断基因组DNA片段的两端连接接头序列,连接产物通过重亚硫酸盐处理将未甲基化修饰的胞嘧啶C转变为尿嘧啶U,进而通过接头序列介导的 PCR 技术将尿嘧啶U转变为胸腺嘧啶T。
应用方向:
WGBS广泛用于各种物种,要求全基因组扫描(不错过关键位点)
- 全基因组甲基化图谱课题
- 标志物筛选课题
- 小规模研究课题
技术优势:
- 应用范围广:适用于所有参考基因组已知物种的甲基化研究;
- 全基因组覆盖:最大限度地获取完整的全基因组甲基化信息,精确绘制甲基化图谱;
- 单碱基分辨率:可精确分析每一个C碱基的甲基化状态。

易基因科技提供全面的DNA甲基化研究整体解决方案,详询易基因:0755-28317900。
参考文献:
Lu Y, Zhou J, Li F, Cao H, Zhang X, Yu D, He Z, Ji H, Lv K, Wu G, Yu M. The Integration of Genome-Wide DNA Methylation and Transcriptomics Identifies the Potential Genes That Regulate the Development of Skeletal Muscles in Ducks. Int J Mol Sci. 2023 Oct 23;24(20) pii: ijms242015476.
相关阅读:
抗性育种 | WGBS等从DNA甲基化揭示杀鲑气单胞菌灭活疫苗对大菱鲆的免疫力
项目文章 | WGBS等揭示SOX30甲基化在非梗阻性无精症中的表观遗传调控机制